Question: How does exercise affect acute and chronic levels of inflammation?
Takeaway: Exercise leads to an acute increase in IL-6, which suppresses production of pro-inflammatory cytokines, including TNF. While this research remains preliminary, it suggests the impact of exercise on IL-6 may explain the beneficial effects of exercise on metabolic and general health.
This 2005 review argues an exercise-induced increase in IL-6 reduces the harmful effects of inflammation and the anti-inflammatory properties of exercise may explain the link between activity and health.
Low-grade, chronic inflammation (i.e., increased systemic levels of C-reactive protein, or CRP, and some cytokines) is associated with a variety of metabolic conditions, including the metabolic syndrome, Type 2 diabetes, and cardiovascular disease (1). Elevated levels of TNF and CRP have more specifically been associated with aging, diabetes, insulin resistance, and general and cause-specific mortality (2). Some of this same research has argued these associations point to a causal link and these same metabolic conditions may be exacerbated or outright caused by inflammation. TNF, specifically, has been found to directly inhibit insulin signaling and increase insulin resistance (3).
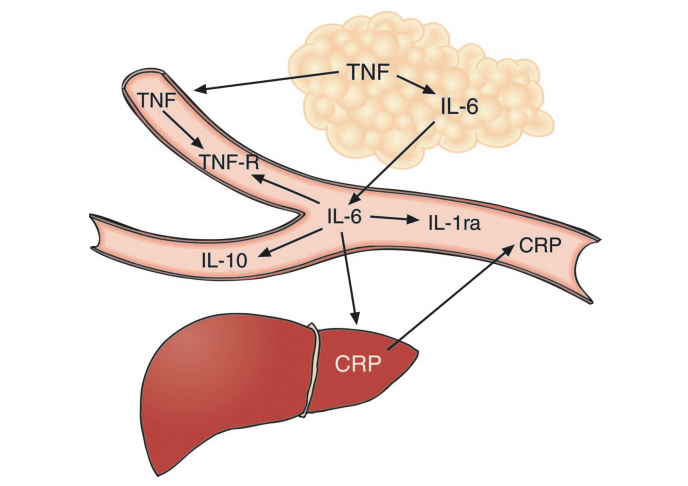
Figure 1: The authors illustrate the relationship between various cytokines. Increased TNF, derived from adipose tissue, leads to increased levels of IL-6 and other cytokines.
Individuals with chronic inflammation often have elevated IL-6 levels, which has led many to conclude IL-6 is a contributor to these same conditions. However, the authors of this review argue IL-6 is merely downstream of TNF and is itself primarily anti-inflammatory in its actions. As shown in the figure below, elevated TNF levels — which are primarily derived from adipose tissue (4) and so are a consequence of increased adiposity — lead to increased IL-6 production. Direct studies of IL-6, however, have consistently shown benefits favoring metabolic health. Rodent, human, and in vitro studies suggest IL-6 enhances lipolysis and increases fat oxidation (5). Genomic studies have found individuals with increased production of TNF and reduced production of IL-6 have an increased risk of diabetes and the metabolic syndrome, suggesting differential roles for TNF and IL-6 (6). This preliminary evidence indicates IL-6 may be beneficial; it may only be present at high levels in metabolically unhealthy individuals because it is downstream of TNF.
Exercise leads to a selective cytokine response. As shown in the figure below, and in direct contrast to the broad inflammatory response associated with sepsis (the classical model of inflammation), the distinguishing cytokine response to exercise is a rapid, dramatic (100-fold) increase in IL-6 followed by smaller elevations in other cytokines downstream of IL-6 (7). As the review authors note, the finding that exercise elevates IL-6 levels without muscle damage is “remarkably consistent” throughout the literature. Intensity, duration, muscle mass recruitment, and training level do seem to moderate IL-6 response (8). Carbohydrate suppresses the exercise-induced IL-6 response, and low muscular glycogen levels increase the response (9). These elevated IL-6 levels — when generated in the context of exercise, without an adipose-derived elevation in TNF levels — inhibit TNF production and reduce TNF levels, as shown in multiple in vitro and mouse studies (10).
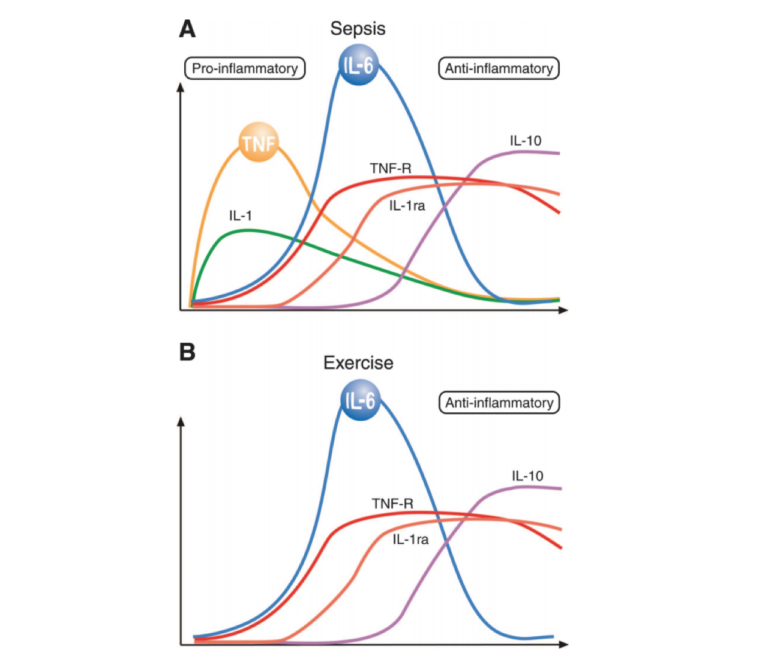
Figure 2: A comparison of the cytokine responses of sepsis and exercise. Exercise does not lead to a significant elevation of TNF; instead, the distinguishing response is a dramatic increase in IL-6 followed by increases in other cytokines downstream of IL-6.
These authors have demonstrated the exercise-induced elevation in IL-6 suppresses TNF activity. In a previous paper (11), subjects were injected with a low dose of E. coli toxin at rest or during exercise. As expected, the toxin led to a two- to three-fold increase in TNF levels in resting subjects; in exercising subjects, however, the TNF response was entirely blunted. Mouse models that overproduce TNF are normalized via exercise (12). Elevated epinephrine levels during exercise may further suppress TNF, though the mechanism of these effects is not yet clear (13).
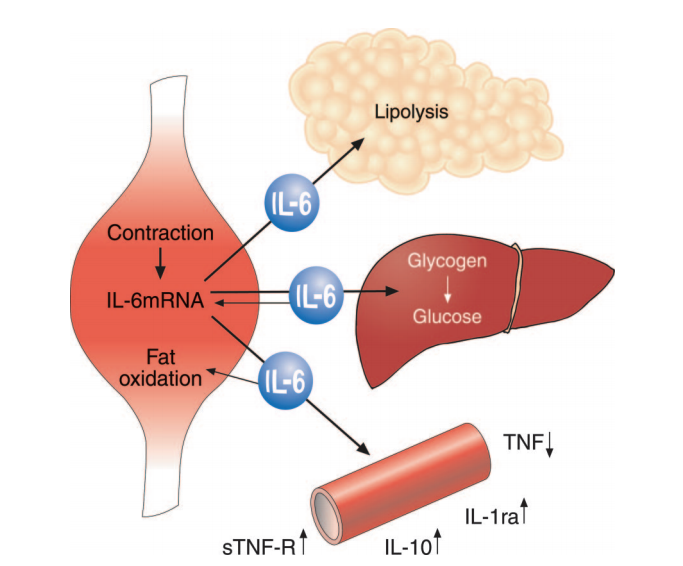
Figure 3: Muscular contraction increases IL-6 production, which stimulates lipolysis, supports glucose regulation during exercise, and suppresses production of pro-inflammatory cytokine TNF.
Taken together, these observations lead the authors to conclude exercise-induced IL-6 elevation produces an anti-inflammatory state by inhibiting production of TNF. These elevated IL-6 levels will simultaneously stimulate lipolysis and fat oxidation, reducing the excess adiposity that contributes to TNF overproduction. The authors argue IL-6 and other cytokines produced as a result of muscular exertion — which they collectively term “myokines” — may thus explain the relationships between increased levels of activity and metabolic health, increased activity and reduced inflammation, and sedentary behavior and metabolic disease (14). Further research could explore how the acute IL-6 response induced by exercise affects chronic inflammation levels and how different exercise programs specifically affect the chronic inflammatory state.
Notes
- The relation of markers of inflammation to the development of glucose disorders in the elderly: The Cardiovascular Health Study; Low-grade systemic inflammation and the development of type 2 diabetes: The atherosclerosis risk in communities study; Elevated levels of acute-phase proteins and plasminogen activator inhibitor-1 predict the development of type 2 diabetes: The insulin resistance atherosclerosis study; Leukocyte count, erythrocyte sedimentation rate, and diabetes incidence in a national sample of US adults; C-reactive protein is an independent predictor of risk for the development of diabetes in the West of Scotland Coronary Prevention Study; Prospective study of C-reactive protein in relation to the development of diabetes and metabolic syndrome in the Mexico City Diabetes Study; Adiponectin and development of type 2 diabetes in the Pima Indian population; White blood-cell count and the risk of impaired fasting glucose or Type II diabetes in middle-aged Japanese men; C-reactive protein, interleukin 6, and risk of developing type 2 diabetes mellitus; High white blood cell count is associated with a worsening of insulin sensitivity and predicts the development of type 2 diabetes; Inflammation: The link between insulin resistance, obesity and diabetes; Inflammation in atherosclerosis; Inflammation and atherosclerosis
- A high plasma concentration of TNF-alpha is associated with dementia in centenarians; Ageing, tumour necrosis factor-alpha (TNF-alpha) and atherosclerosis; Association of circulating TNF-alpha and IL-6 with ageing and parkinsonism; Exercise induces recruitment of lymphocytes with an activated phenotype and short telomere lengths in young and elderly humans; Advancing age and insulin resistance: role of plasma tumor necrosis factor-α; Reactive protein and other markers of inflammation in the prediction of cardiovascular disease in women; Plasma concentration of interleukin-6 and the risk of future myocardial infarction among apparently healthy men; The expression of TNF alpha by human muscle. Relationship to insulin resistance; Role of cytokines in inducing hyperlipidemia; Relationship between serum tumor necrosis factor-alpha and insulin resistance in obese men with Type 2 diabetes mellitus; Elevated serum tumor necrosis factor-alpha concentrations and bioactivity in Type 2 diabetics and patients with android type obesity; Pro-inflammatory cytokines and adipose tissue; Adipose expression of tumor necrosis factor-alpha: Direct role in obesity-linked insulin resistance
- Effects of tumor necrosis factor-alpha on insulin action in cultured human muscle cells; Acute impairment of insulin-mediated capillary recruitment and glucose uptake in rat skeletal muscle in vivo by TNF-alpha; Protection from obesity-induced insulin resistance in mice lacking TNF-alpha function; Neutralization of tumor necrosis factor reverses age-induced impairment of insulin responsiveness in skeletal muscle of Sprague-Dawley rats; Mechanisms of TNF-alpha-induced insulin resistance; IRS-1-mediated inhibition of insulin receptor tyrosine kinase activity in TNF-alpha- and obesity-induced insulin resistance; Tumor necrosis factor (TNF)-alpha inhibits insulin signaling through stimulation of the p55 TNF receptor and activation of sphingomyelinase; Inhibition of proteasome activity blocks the ability of TNFα to down-regulate Gi proteins and stimulate lipolysis; Tumor necrosis factor alpha stimulates lipolysis in adipocytes by decreasing Gi protein concentrations; Tumor necrosis factor increases the rate of lipolysis in primary cultures of adipocytes without altering levels of hormone-sensitive lipase; Overexpression of perilipin A and B blocks the ability of tumor necrosis factor alpha to increase lipolysis in 3T3–L1 adipocytes; BRL 49653 blocks the lipolytic actions of tumor necrosis factor-alpha: a potential new insulin-sensitizing mechanism for thiazolidinediones
- Pro-inflammatory cytokines and adipose tissue; Adipose expression of tumor necrosis factor-alpha: Direct role in obesity-linked insulin resistance
- Acute IL-6 treatment increases fatty acid turnover in elderly humans in vivo and in tissue culture in vitro: evidence that IL-6 acts independently of lipolytic hormones; Interleukin-6-deficient mice develop mature-onset obesity; Interleukin-6 stimulates lipolysis and fat oxidation in humans; Cytokine regulation of skeletal muscle fatty acid metabolism: effect of interleukin-6 and tumor necrosis factor-α; Interleukin-6 stimulates hepatic triglyceride secretion in rats; Human breast adipocytes express interleukin-6 (IL-6) and its receptor system: increased IL-6 production by beta-adrenergic activation and effects of IL-6 on adipocyte function; Acute IL-6 treatment increases fatty acid turnover in elderly humans in vivo and in tissue culture in vitro: evidence that IL-6 acts independently of lipolytic hormones; Endocrinologic and metabolic effects of interleukin-6 in humans
- Promoter polymorphisms of the TNF-alpha (G-308A) and IL-6 (C-174G) genes predict the conversion from impaired glucose tolerance to type 2 diabetes: The Finnish Diabetes Prevention Study
- Muscle-derived interleukin-6: mechanisms for activation and possible biological roles; Exercise and the immune system: regulation, integration and adaptation; Searching for the exercise factor: is IL-6 a candidate; Muscle-derived interleukin-6: possible biological effects; Systemic inflammatory response to exhaustive exercise. Cytokine kinetics; Some aspects of the acute phase response after a marathon race, and the effects of glutamine supplementation; Endurance run increases circulating IL-6 and IL-1ra but downregulates ex vivo TNF-α and IL-1β production; Leukemia inhibitory factor, Interleukin 6, and other cytokines using the GP130 transducing receptor: roles in inflammation and injury; Xanthine oxidase in human skeletal muscle following eccentric exercise: a role of inflammation; Carbohydrate and the cytokine response to 2.5 hours of running; Lymphocytes and NK cell activity during repeated bouts of maximal exercise; Effects of mode and carbohydrate on the granulocyte and monocyte response to intensive prolonged exercise; Influence of mode and carbohydrate on the cytokine response to heavy exertion; The cytokine response to strenuous exercise; A trauma-like elevation in plasma cytokines in humans in response to treadmill running; Chemokines are elevated in plasma after strenuous exercise in humans; Pro- and anti-inflammatory cytokine balance in strenuous exercise in humans; Evidence that IL-6 is produced in skeletal muscle during prolonged running; Physical activity and plasma interleukin-6 in humans—effect of intensity of exercise; Prolonged submaximal eccentric exercise is associated with increased levels of plasma IL-6; Carbohydrate ingestion attenuates the increase in plasma interleukin-6, but not skeletal muscle interleukin-6 mRNA, during exercise in humans; Circulating monocytes are not the source of elevations in plasma IL-6 and TNF-α levels after prolonged running; Interleukin-6 production in contracting human skeletal muscle is influenced by pre-exercise muscle glycogen content; IL-6 and TNF-α expression in, and release from, contracting human skeletal muscle; Plasma interleukin-6 during strenuous exercise: Role of adrenaline; Production of interleukin-6 in contracting human skeletal muscles can account for the exercise-induced increase in plasma interleukin-6; Circulating cytokines and hormones with immunosuppressive but neutrophil-priming potentials rise after endurance exercise in humans; The effects of n-3 PUFA on the cytokine response to strenuous exercise
- Does the aging skeletal muscle maintain its endocrine function?; Supplementation with vitamins C and E inhibits the release of interleukin-6 from contracting human skeletal muscle
- Transcriptional activation of the IL-6 gene in human contracting skeletal muscle: influence of muscle glycogen content; Carbohydrate and the cytokine response to 2.5 hours of running; Influence of mode and carbohydrate on the cytokine response to heavy exertion; Interleukin-6 production in contracting human skeletal muscle is influenced by pre-exercise muscle glycogen content
- Anti-gamma interferon and anti-interleukin-6 antibodies affect staphylococcal enterotoxin B-induced weight loss, hypoglycemia, and cytokine release in d-galactosamine-sensitized and unsensitized mice; T cell activation-associated hepatic injury: mediation by tumor necrosis factors and protection by interleukin 6; Correlations and interactions in the production of interleukin-6 (IL-6), IL-1, and tumor necrosis factor (TNF) in human blood mononuclear cells: IL-6 suppresses IL-1 and TNF
- Exercise and IL-6 infusion inhibit endotoxin-induced TNF-α production in humans
- Exercise normalises overexpression of TNF-alpha in knockout mice
- Epinephrine inhibits tumor necrosis factor-alpha and potentiates interleukin 10 production during human endotoxemia
- Is physical activity or physical fitness more important in defining health benefits?; Exercise-based rehabilitation for coronary heart disease; Exercise training meta-analysis of trials in patients with chronic heart failure (ExTraMATCH); Effects of exercise on glycemic control and body mass in type 2 diabetes mellitus: a meta-analysis of controlled clinical trials; Relationship between physical activity and inflammation among apparently healthy middle-aged and older US adults; The acute phase response and exercise: court and field sports; Association between physical activity and markers of inflammation in a healthy elderly population; Inflammatory markers and exercise: Differences related to exercise type; Reduction of the plasma concentration of C-reactive protein following nine months of endurance training; Long-term exercise and atherogenic activity of blood mononuclear cells in persons at risk of developing ischemic heart disease; Cross-sectional and prospective relationships of interleukin-6 and C-reactive protein with physical performance in elderly persons: MacArthur studies of successful aging; Physical activity and hemostatic and inflammatory variables in elderly men
Comments on The Anti-Inflammatory Effect of Exercise
Studies like this definitely help to clarify why a stressor like exercise can provide positive benefits while other stressors can largely be negative to the organism.
The Anti-Inflammatory Effect of Exercise
1